KDM5C Antibodies
Background
The KDM5C gene encodes a histone delysinase, which is mainly present in the cell nucleus of animals. This protein can specifically remove the trimethylation modification of lysine at position 4 of histone H3, thereby regulating chromatin structure and gene transcriptional activity, and plays a key role in neural development and X chromosome dose compensation. Because this gene is located on the X chromosome and shows haploid deficiency in male individuals, its mutation often leads to X-linked intellectual disability syndrome. In 2007, the team led by scientist R. J. Klose clarified the mechanism of its demethylase. This discovery not only deepened the understanding of the epigenetic regulatory network but also provided an important model for studying the association between chromatin remodeling and cognitive dysfunction, promoting the cross-integration of epigenetics and neurodevelopmental disease research.
Structure of KDM5C
KDM5C is a histone delysinase with a molecular weight of approximately 180 kDa. This protein shows significant structural differences among different species, especially in the sequence conservation of its catalytic domain and PHD domain, which demonstrates evolutionary divergence.
| Species | Human | Mouse | Zebrafish | Fruit fly |
| Molecular Weight (kDa) | 180.2 | 178.9 | 175.3 | 165.8 |
| Primary Structural Differences | Contains the JmjC catalytic domain and multiple PHD zinc fingers | JmjN domain has three amino acid replacement | Lack of a PHD domain | Only the core catalytic unit is retained |
KDM5C is composed of 1,560 amino acids, and its three-dimensional structure contains a characteristic JmjC catalytic domain and multiple PHD zinc finger modules. The active center of this enzyme achieves demethylation of histone H3K4me3/me2 through the synergistic action of Fe(II) and α -ketoglutaric acid. The substrate recognition specificity is determined by the reading ability of the third PHD zinc finger to H3K4me0, while the ARID domain at the N-terminal is responsible for chromatin binding. This multi-domain collaborative assembly enables KDM5C to precisely regulate gene transcriptional activity.
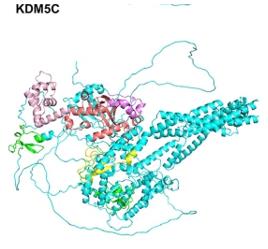 Fig. 1 KDM5C structure.1
Fig. 1 KDM5C structure.1
Key structural properties of KDM5C:
- Contains the conserved JmjC catalytic domain
- Protein-protein interaction modules composed of multiple PHD zinc fingers
- Dependent on the active centers of Fe(II) and α -ketoglutaric acid
Functions of KDM5C
The core function of the KDM5C gene is to regulate the transcriptional activity of genes. However, it is also involved in a variety of biological processes, including cell differentiation, tumorigenesis and neural development.
| Function | Description |
| Transcriptional inhibition | Inhibit the transcription initiation of target genes by catalyzing the demethylation of histone H3K4me3/me2. |
| Regulation of cell differentiation | Maintain the "transcription-ready" state of a specific genome in embryonic stem cells and coordinate the differentiation process. |
| Tumor suppression | The inactivation mutations can lead to multiple abnormal activation of oncogene, is associated with kidney cancer and breast cancer. |
| Intellectual development | By adjusting the synapses related gene chromatin state, affect cognitive function development. |
| X chromosome dose compensation | As an X-linked gene, it participates in the balanced regulation of X chromosome activity in male individuals. |
The kinetic characteristics of this enzyme are manifested as its specific recognition of H3K4me3, and its catalytic efficiency is regulated by the allosteric changes of the PHD zinc finger and the substrate. This multiple regulatory mechanism enables it to precisely respond to intracellular signal transduction pathways.
Applications of KDM5C and KDM5C Antibody in Literature
1. Karwacki-Neisius, Violetta, et al. "WNT signalling control by KDM5C during development affects cognition." Nature 627.8004 (2024): 594-603. https://doi.org/10.1038/s41586-024-07067-y
The article indicates that mutations in the KDM5C gene lead to intellectual disability by disrupting the WNT signaling during the critical period of neural development. Studies have confirmed that briefly regulating the WNT pathway during this window period can successfully salvage the cognitive behavioral deficits of model mice.
2. Chang, Soojeong, Sujin Yim, and Hyunsung Park. "The cancer driver genes IDH1/2, JARID1C/KDM5C, and UTX/KDM6A: crosstalk between histone demethylation and hypoxic reprogramming in cancer metabolism." Experimental & molecular medicine 51.6 (2019): 1-17. https://doi.org/10.1038/s12276-019-0230-6
The article indicates that tumor metabolic reprogramming can produce metabolites that inhibit the activity of histone demethylases such as KDM5C, thereby driving carcinogenesis. This gene family plays distinct roles in different cancers and possesses both oncogene and tumor suppressor gene functions.
3. Liu, Huadie, et al. "The histone demethylase KDM5C controls female bone mass by promoting energy metabolism in osteoclasts." Science Advances 9.14 (2023): eadg0731. https://doi.org/10.1126/sciadv.adg0731
Research has found that the epigenetic factor KDM5C on the X chromosome is the key internal factor that makes women more prone to osteoporosis. It promotes bone resorption by enhancing the energy metabolism of osteoclasts. Inhibiting KDM5C can effectively slow down bone loss in women, providing a new target for the treatment of osteoporosis in women.
4. Müller, Marvin, et al. "KDM5C and KDM5D mutations have different consequences in clear cell renal cell carcinoma cells." Communications biology 8.1 (2025): 244. https://doi.org/10.1038/s42003-025-07695-8
Research has found that in male clear cell renal cell carcinoma, the functional mutation of KDM5C needs to be accompanied by the loss of the Y chromosome (carrying the homologous gene KDM5D) to synergistically promote tumor development, indicating that the functions of the two are not simply redundant.
5. Xiao, Qiong, et al. "Regulation of KDM5C stability and enhancer reprogramming in breast cancer." Cell Death & Disease 13.10 (2022): 843. https://doi.org/10.1038/s41419-022-05296-5
Research has revealed that TRIM11 drives the malignant progression of breast cancer by ubiquitinating and degrading KDM5C, altering the H3K4me3 level on the enhancer. The expressions of the two were negatively correlated in the patient's tissues.
Creative Biolabs: KDM5C Antibodies for Research
Creative Biolabs specializes in the production of high-quality KDM5C antibodies for research and industrial applications. Our portfolio includes monoclonal antibodies tailored for ELISA, Flow Cytometry, Western blot, immunohistochemistry, and other diagnostic methodologies.
- Custom KDM5C Antibody Development: Tailor-made solutions to meet specific research requirements.
- Bulk Production: Large-scale antibody manufacturing for industry partners.
- Technical Support: Expert consultation for protocol optimization and troubleshooting.
- Aliquoting Services: Conveniently sized aliquots for long-term storage and consistent experimental outcomes.
For more details on our KDM5C antibodies, custom preparations, or technical support, contact us at email.
Reference
- Li, Chang-Yun, et al. "KDM5 family as therapeutic targets in breast cancer: Pathogenesis and therapeutic opportunities and challenges." Molecular cancer 23.1 (2024): 109. https://doi.org/10.1186/s12943-024-02011-0
Anti-KDM5C antibodies
 Loading...
Loading...




Hot products 
-
Rat Anti-CCR2 Recombinant Antibody (475301) (CBMAB-C1338-LY)

-
Mouse Anti-ACTB Recombinant Antibody (V2-179553) (CBMAB-A0870-YC)

-
Mouse Anti-AGO2 Recombinant Antibody (V2-634169) (CBMAB-AP203LY)

-
Human Anti-SARS-CoV-2 Spike Recombinant Antibody (CBC05) (CBMAB-CR005LY)

-
Rabbit Anti-ALDOA Recombinant Antibody (D73H4) (CBMAB-A2314-YC)

-
Mouse Anti-COL1A2 Recombinant Antibody (CF108) (V2LY-1206-LY626)

-
Mouse Anti-AMOT Recombinant Antibody (CBYC-A564) (CBMAB-A2552-YC)

-
Mouse Anti-Acetyl-α-Tubulin (Lys40) Recombinant Antibody (V2-623485) (CBMAB-CP2897-LY)

-
Mouse Anti-APP Recombinant Antibody (DE2B4) (CBMAB-1122-CN)

-
Rat Anti-CD63 Recombinant Antibody (7G4.2E8) (CBMAB-C8725-LY)

-
Mouse Anti-CCND2 Recombinant Antibody (DCS-3) (CBMAB-G1318-LY)

-
Mouse Anti-AGK Recombinant Antibody (V2-258056) (CBMAB-M0989-FY)

-
Mouse Anti-4-Hydroxynonenal Recombinant Antibody (V2-502280) (CBMAB-C1055-CN)

-
Mouse Anti-AP4E1 Recombinant Antibody (32) (CBMAB-A2996-YC)

-
Mouse Anti-AKT1 (Phosphorylated S473) Recombinant Antibody (V2-505430) (PTM-CBMAB-0067LY)

-
Mouse Anti-CCDC25 Recombinant Antibody (CBLC132-LY) (CBMAB-C9786-LY)

-
Mouse Anti-CD2AP Recombinant Antibody (BR083) (CBMAB-BR083LY)

-
Mouse Anti-ADAM12 Recombinant Antibody (V2-179752) (CBMAB-A1114-YC)

-
Mouse Anti-CD59 Recombinant Antibody (CBXC-2097) (CBMAB-C4421-CQ)

-
Rabbit Anti-Acetyl-Histone H3 (Lys36) Recombinant Antibody (V2-623395) (CBMAB-CP0994-LY)

- AActivation
- AGAgonist
- APApoptosis
- BBlocking
- BABioassay
- BIBioimaging
- CImmunohistochemistry-Frozen Sections
- CIChromatin Immunoprecipitation
- CTCytotoxicity
- CSCostimulation
- DDepletion
- DBDot Blot
- EELISA
- ECELISA(Cap)
- EDELISA(Det)
- ESELISpot
- EMElectron Microscopy
- FFlow Cytometry
- FNFunction Assay
- GSGel Supershift
- IInhibition
- IAEnzyme Immunoassay
- ICImmunocytochemistry
- IDImmunodiffusion
- IEImmunoelectrophoresis
- IFImmunofluorescence
- IGImmunochromatography
- IHImmunohistochemistry
- IMImmunomicroscopy
- IOImmunoassay
- IPImmunoprecipitation
- ISIntracellular Staining for Flow Cytometry
- LALuminex Assay
- LFLateral Flow Immunoassay
- MMicroarray
- MCMass Cytometry/CyTOF
- MDMeDIP
- MSElectrophoretic Mobility Shift Assay
- NNeutralization
- PImmunohistologyp-Paraffin Sections
- PAPeptide Array
- PEPeptide ELISA
- PLProximity Ligation Assay
- RRadioimmunoassay
- SStimulation
- SESandwich ELISA
- SHIn situ hybridization
- TCTissue Culture
- WBWestern Blot
